染色体
定義-染色体とは何ですか?
細胞の遺伝子構成は、DNA(デオキシリボ核酸)とその塩基(アデニン、チミン、グアニン、シトシン)の形で保存されています。すべての真核細胞(動物、植物、真菌)では、これは染色体の形で細胞核に存在します。染色体は、特定のタンパク質にリンクされた単一のコヒーレントDNA分子で構成されています。
染色体という名前はギリシャ語に由来し、大まかに「カラーボディ」と翻訳することができます。この名前は、細胞学の歴史の非常に早い時期(1888)に、科学者が特殊な塩基性染料を使用してそれらを染色し、光学顕微鏡でそれらを識別することに成功したという事実に由来しています。しかし、それらは、染色体が特に密集している(凝縮している)細胞周期の特定の時点である有糸分裂(生殖細胞では減数分裂)でのみ実際に見られます。

染色体はどのように作られていますか?
細胞のDNA二重らせん全体、つまり約3.4 x 109の塩基対が互いに結合している場合、長さは1メートルを超えます。追加されたすべての染色体の全長はわずか約115µmです。この長さの違いは、染色体の非常にコンパクトな構造によって説明されます。この構造では、DNAが非常に特殊な方法で数回巻かれたりらせん状に巻かれたりします。
タンパク質の特殊な形態であるヒストンは、これに重要な役割を果たします。 H1、H2A、H2B、H3、H4の合計5つの異なるヒストンがあります。最後の4つのヒストンのうちの2つが組み合わさって円筒形の構造、オクタマーを形成し、その周りに二重らせんが約2回巻かれます(=スーパーらせん)。 H1は、この構造を安定させるために、この構造に付着します。
このDNA、オクタマー、H1の複合体は、ヌクレオソームと呼ばれます。これらのヌクレオソームのいくつかは、現在、比較的短い間隔(10〜60塩基対)で「真珠のストリングのように」前後に並んでいます。染色体間のセクションはスペーサーDNAと呼ばれます。個々のヌクレオソームはH1を介して再び接触し、さらにらせん状になり、圧縮も生じます。
結果として生じる鎖は、順番に、ヘルトンとしても知られている酸性の非ヒストンタンパク質で作られたバックボーンによって安定化されたループにあります。これらのループは、タンパク質によって安定化されたスパイラルに存在し、圧縮の最終段階になります。ただし、この高度な圧縮は、有糸分裂中の細胞分裂の状況でのみ発生します。
このフェーズでは、2つの染色分体で構成される染色体の特徴的な形状も確認できます。これらがつながっている場所をセントロメアと呼びます。これは、各中期染色体を、pアームおよびqアームとも呼ばれる2つの短いアームと2つの長いアームに分割します。
セントロメアが染色体のほぼ中央にある場合、それが完全にアクロセントリック染色体の一方の端にある場合、それはメタセントリック染色体と呼ばれます。その間にあるものはサブメタセントリック染色体と呼ばれます。光学顕微鏡ですでに確認できるこれらの違いは、長さとともに、染色体の初期分類を可能にします。
テロメアとは何ですか?
テロメアは、繰り返し配列を持つ染色体の末端です(TTAGGG)。これらは関連情報を持っていませんが、より関連性の高いDNAセクションの損失を防ぐのに役立ちます。細胞分裂のたびに、染色体の一部がDNA複製のメカニズムによって失われます。
したがって、テロメアは、ある意味で、細胞が分裂することによって重要な情報を失うポイントを遅らせるバッファーです。細胞のテロメアの長さが4,000塩基対未満の場合、プログラムされた細胞死(アポトーシス)が開始されます。これにより、欠陥のある遺伝物質が生物に広がるのを防ぎます。いくつかの細胞はテロメラーゼ、すなわちテロメアを再び伸ばすことができる酵素を持っています。
他のすべての細胞が発生する幹細胞に加えて、これらは生殖細胞と免疫系の特定の細胞です。さらに、テロメラーゼは癌細胞にも見られます。そのため、この文脈で細胞の不死化について話します。
ここでトピックに関するすべてを読んでください:テロメア-解剖学、機能および病気
クロマチンとは何ですか?
クロマチンとは、塩基で染色できる細胞核の全内容物を指します。したがって、この用語には、DNAに加えて、特定のタンパク質、たとえばヒストンやヘルトン(構造を参照)、および特定のRNAフラグメント(hnおよびsnRNA)も含まれます。
細胞周期の段階または遺伝的活動に応じて、この材料はさまざまな密度で入手できます。より密度の高い形態はヘテロクロマチンと呼ばれます。したがって、理解しやすくするために、それを「貯蔵形態」と見なし、ここでも構成的ヘテロクロマチンと通性的ヘテロクロマチンを区別することができます。
構成的ヘテロクロマチンは最も密度の高い形態であり、細胞周期のすべての段階で最高レベルの凝縮で存在します。ヒトゲノムの約6.5%を占め、主にセントロメアと染色体アーム(テロメア)の端の近くにわずかに位置していますが、他の場所(主に染色体1、9、16、19、Y)にも位置しています。 。さらに、構成的ヘテロクロマチンのほとんどは、核膜の近く、すなわち細胞核の端に位置しています。中央のスペースは、アクティブなクロマチンであるユークロマチンのために予約されています。
通性ヘテロクロマチンは少し密度が低く、必要に応じて、または開発の段階に応じて、活性化および非活性化することができます。この良い例は、女性の核型の2番目のX染色体です。 1つのX染色体は基本的に細胞が生き残るのに十分であり、最終的には男性にも十分であるため、2つのうちの1つは胚期に非活性化されます。非活性化されたX染色体はバー小体として知られています。
細胞分裂の間だけ、有糸分裂の文脈で、それは完全に凝縮し、それによって中期で最高の圧縮に達します。ただし、異なる遺伝子は頻繁に異なる方法で読み取られるため(結局のところ、すべてのタンパク質が常に同じ量で必要とされるわけではありません)、ここでもアクティブなユークロマチンと非アクティブなユークロマチンが区別されます。
これについてもっと読む:クロマチン
一倍体染色体
一倍体(ギリシャのハプロス=単一)は、細胞のすべての染色体が個別に存在することを意味します。つまり、通常の場合のようにペア(二倍体)ではありません。これはすべての卵子と精子細胞の自然な状態であり、最初の減数分裂の過程で2つの同一の染色分体が最初に分離されるのではなく、染色体のすべてのペアが最初に分離されます。
その結果、最初の減数分裂後、ヒトの娘細胞は通常の46染色体ではなく、23染色体しか持たず、これは染色体の半数体セットの半分に相当します。これらの娘細胞はまだ2つの染色体からなる各染色体の同一のコピーを持っているので、2つの染色分体が互いに分離されている2番目の減数分裂が必要です。
多糸染色体
多糸染色体は、多数の遺伝的に同一の染色分体で構成された染色体です。このような染色体は、低倍率でも見やすいため、巨大染色体と呼ばれることもあります。このための前提条件は、細胞核内の染色体が細胞分裂を起こさずに数回増殖する核内倍加です。
染色体の機能は何ですか?
私たちのゲノムの組織単位としての染色体は、主に、細胞分裂中に倍加されたゲノムが娘細胞間で均等に分布することを保証するのに役立ちます。これを行うには、細胞分裂または細胞周期のメカニズムを詳しく調べる価値があります。
細胞は細胞周期の大部分を間期に費やします。これは、細胞がすぐに分裂しようとしない期間全体を意味します。これは次にG1、S、G2期に分けられます。
G1期(ギャップのようにG、すなわちギャップ)は細胞分裂の直後に続きます。ここで細胞は再びサイズが大きくなり、一般的な代謝機能を実行します。
ここから、G0期に切り替えることもできます。これは、分裂することができなくなった段階に変化し、通常の場合、非常に特定の機能(細胞分化)を果たすために大きく変化することを意味します。これらのタスクを実行するために、非常に特定の遺伝子がより強く読み取られ、他の遺伝子はより少なくまたはまったく読み取られません。
DNAのセグメントが長期間必要とされない場合、それは多くの場合、長期間密に詰まっている染色体の部分にあります(クロマチンを参照)。一方で、これはスペースを節約する目的を持っていますが、遺伝子調節の他のメカニズムに加えて、誤って読み取られることに対する追加の保護でもあります。しかし、非常に特殊な条件下では、G0期から分化した細胞が再びサイクルに入る可能性があることも観察されています。
G1期の後にS期、つまり新しいDNAが合成される期(DNA複製)が続きます。ここでは、DNA全体が最も緩い形である必要があります。つまり、すべての染色体が完全にほどかれています(構造を参照)。
合成段階の終わりに、遺伝物質全体が細胞内に重複して存在します。コピーはまだセントロメアを介して元の染色体に付着しているため(構造を参照)、染色体の重複については話しません。
各染色体は現在、1つではなく2つの染色分体で構成されているため、後で有糸分裂中に特徴的なX字型をとることができます(厳密に言えば、X字型はメタセントリック染色体にのみ適用されます)。次のG2期では、細胞分裂の即時準備が行われます。これには、必要に応じて修復できるレプリケーションエラーとストランドブレークの詳細なチェックも含まれます。
細胞分裂には、基本的に有糸分裂と減数分裂の2種類があります。生殖細胞を除いて、生物のすべての細胞は有糸分裂によって生じます。その唯一の仕事は、2つの遺伝的に同一の娘細胞の形成です。
一方、減数分裂には、遺伝的に異なる細胞を生成する目的があります。
最初のステップでは、対応する(相同な)が同一ではない染色体が分割されます。次のステップでのみ、2つの同一の染色分体からなる染色体が分離され、それぞれ2つの娘細胞に再び分配されます。その結果、最終的に、異なる遺伝物質を持つ4つの生殖細胞が1つの芽球から発生します。
染色体の形態と構造は、両方のメカニズムに不可欠です。特別な「タンパク質糸」、いわゆる紡錘体装置は、高度に凝縮された染色体に付着し、中央面(赤道面)から細かく制御されたプロセスで染色体を引き出します。均等な分布を確保するために、1つの周りのセルの反対の極に。染色体の微細構造のわずかな変化でさえ、ここで深刻な結果をもたらす可能性があります。
すべての哺乳動物において、性染色体XとYの比率も子孫の性を決定します。基本的に、それはすべて、卵細胞と結合する精子がX染色体を持っているかY染色体を持っているかによって異なります。両方の形態の精子は常に正確に同じ程度に生成されるので、確率は常に両方の性別でバランスが取れています。このランダムシステムは、たとえば気温などの環境要因の場合よりも、より均一な性別分布を保証します。
トピックの詳細をご覧ください:細胞核分裂
遺伝子構成は染色体を介してどのように受け継がれますか?
今日、私たちは形質がDNAの形で細胞に保存されている遺伝子を介して受け継がれていることを知っています。これらは順番に46の染色体に分割され、その上に25,000〜30000のヒト遺伝子が分布しています。
表現型と呼ばれる特性自体に加えて、遺伝子型と呼ばれる遺伝的同等物もあります。遺伝子が染色体上にある場所は遺伝子座と呼ばれます。人間はすべての染色体が2倍あるため、すべての遺伝子も2回発生します。これに対する唯一の例外は、男性のX染色体遺伝子です。これは、Y染色体がX染色体上にある遺伝情報の一部しか持っていないためです。
同じ遺伝子座にある異なる遺伝子は対立遺伝子と呼ばれます。多くの場合、1つの遺伝子座に3つ以上の異なる対立遺伝子があります。次に、ポリモーフィズムについて話します。このような対立遺伝子は、単に無害な変異体(正常な変異体)である可能性がありますが、遺伝性疾患の引き金となる可能性のある病理学的変異でもあります。
単一遺伝子の突然変異が表現型を変えるのに十分である場合、単一遺伝子またはメンデルの法則について話します。ただし、継承可能な特性の多くは、いくつかの相互作用する遺伝子を介して継承されるため、研究がはるかに困難です。
メンデルの法則では、母親と父親がそれぞれ2つの遺伝子の1つを子供に渡すため、次世代では常に4つの可能な組み合わせがあり、これらは1つのプロパティに関して同じである可能性があります。個体の両方の対立遺伝子が表現型に同じ影響を与える場合、個体はこの特徴に関してホモ接合性であり、それに応じて特徴は完全に発現される。
ヘテロ接合体には、異なる方法で相互作用できる2つの異なる対立遺伝子があります。ある対立遺伝子が別の対立遺伝子よりも優性である場合、その発現が完全に抑制され、優性形質が表現型に現れます。抑制された対立遺伝子は劣性と呼ばれます。
共優性遺伝の場合、両方の対立遺伝子は互いに影響を受けずに自分自身を表現することができますが、中間遺伝の場合、両方の特性が混在しています。この良い例は、AとBが互いに優勢であるが、0が互いに優勢であるAB0血液型システムです。
人間の染色体の正常なセットは何ですか?
人間の細胞には、22の性非依存性染色体ペア(常染色体)と2つの性染色体(ゴノソーム)があるため、合計46の染色体が1セットの染色体を構成します。
常染色体は通常ペアで来ます。ペアの染色体は、遺伝子の形状と配列が類似しているため、相同と呼ばれます。女性の2つのX染色体も相同ですが、男性はX染色体とY染色体を持っています。これらは、存在する遺伝子の形態と数が異なり、相同性について話すことができなくなります。
生殖細胞、すなわち卵子と精子細胞は、減数分裂のために染色体セットの半分、つまり22個の常染色体とそれぞれ1個のゴノソームしか持っていません。生殖細胞は受精中に融合し、時にはセグメント全体を交換する(クロスオーバー)ため、染色体の新しい組み合わせ(組換え)が作成されます。すべての染色体は一緒に核型と呼ばれ、いくつかの例外(染色体異常を参照)を除いて、同性のすべての個体で同一です。
ここでは、トピックに関するすべてを見つけることができます:有糸分裂-簡単に説明します!
なぜ染色体のペアが常にあるのですか?
基本的に、この質問は一文で答えることができます:それは有益であることが示されているからです。染色体ペアの存在と組換えの原理は、有性生殖の観点から遺伝に不可欠です。このようにして、まったく新しい個体が偶然に2人の個体の遺伝物質から出現する可能性があります。
このシステムは、種内の特性の多様性を大幅に増加させ、突然変異と選択によってのみ可能であるよりもはるかに速く、より柔軟に変化する環境条件に適応できることを保証します。
染色体の二重のセットには保護効果もあります。遺伝子の突然変異が機能の失敗につながる場合でも、2番目の染色体には一種の「バックアップコピー」があります。これは、特に変異した対立遺伝子が優勢である場合、生物が機能不全を補うのに必ずしも十分ではありませんが、それはその可能性を高めます。さらに、この方法では、突然変異がすべての子孫に自動的に渡されるわけではなく、その結果、種が過度に根本的な突然変異から保護されます。
染色体変異とは何ですか?
遺伝的欠陥は、電離放射線(X線など)、化学物質(タバコの煙に含まれるベンゾピレンなど)、特定のウイルス(HPウイルスなど)から発生する可能性があります。また、偶然に発生する可能性もあります。多くの場合、その開発にはいくつかの要因が関係しています。原則として、このような変化はすべての体組織で発生する可能性がありますが、実際的な理由から、分析は通常、リンパ球(特殊なタイプの免疫細胞)、線維芽細胞(結合組織細胞)、および骨髄細胞に限定されます。
染色体変異は、個々の染色体の主要な構造変化です。一方、染色体全体の欠如または追加は、ゲノムまたは倍数性の突然変異であり、遺伝子突然変異という用語は、遺伝子内の比較的小さな変化を指します。染色体異常(ラテン語の異常=逸脱する)という用語はやや広範で、光学顕微鏡で検出できるすべての変化が含まれます。
突然変異は非常に異なる影響を与える可能性があります。
- サイレント突然変異、つまり変化が個人またはその子孫に影響を及ぼさない突然変異は、染色体異常にはかなり異例であり、遺伝子または点突然変異の領域でより頻繁に見られます。
- 突然変異が誤って折りたたまれ、したがって機能のないタンパク質をもたらすか、またはタンパク質をまったくもたらさない場合、機能喪失突然変異について話す。
- いわゆる機能獲得型変異は、まったく新しい効果が生じるように、効果のタイプまたは生成されるタンパク質の量を変化させます。一方で、これは進化のための、したがって種の生存または新種の出現のための重要なメカニズムですが、他方では、フィラデルフィア染色体の場合のように、それはまた決定的な貢献をすることができます癌細胞の発生。
さまざまな形態の染色体異常の中で最もよく知られているのは、おそらく数値異常であり、個々の染色体は1回だけ(一染色体)、さらには3回(トリソミー)存在します。
これが単一の染色体にのみ当てはまる場合、それは異数性と呼ばれ、染色体セット全体が倍数性(3倍および4倍)の影響を受けます。ほとんどの場合、この偏在は、細胞分裂(有糸分裂)中の染色体の非分離(非分離)を介した生殖細胞の発達の過程で発生します。これは、娘細胞上の染色体の不均一な分布につながり、したがって、子供の数値異常につながります。
非性染色体(=常染色体)の一染色体は生命と両立しないため、生きている子供には発生しません。トリソミー13、18、21を除いて、常染色体トリソミーはほとんどの場合自然流産につながります。
いずれにせよ、目立たないこともある性染色体の異常とは対照的に、常に深刻な臨床症状があり、原則として、多かれ少なかれ顕著な外部異常(異形)があります。
このような偏在は、有糸分裂細胞分裂(生殖細胞を除くすべての細胞)を伴う後年にも発生する可能性があります。影響を受けた細胞に加えて変化していない細胞があるので、体細胞モザイクについて話します。体細胞(ギリシャ語の体=体)とは、生殖細胞ではないすべての細胞を意味します。体の細胞のごく一部だけが影響を受けるので、症状は通常はるかに穏やかです。したがって、モザイクタイプは長い間検出されないままになることがよくあります。
ここでは、トピックに関するすべてを見つけることができます:染色体変異
染色体異常とは何ですか?
構造的染色体異常は基本的に染色体突然変異の定義に対応します(上記参照)。遺伝物質の量が同じままで、単純に異なって分布している場合、バランスの取れた異常について話します。
これは多くの場合、乗換え、つまり染色体セグメントの別の染色体への移動を介して行われます。それが2つの染色体間の交換である場合、1つは相互転置について話します。タンパク質を生産するのに必要なゲノムは約2%しかないため、そのような遺伝子がブレークポイントにあり、その機能を失ったり、機能が損なわれたりする可能性は非常に低くなります。したがって、このようなバランスの取れた収差は見過ごされがちで、数世代にわたって受け継がれます。
しかし、これは生殖細胞の発達中に染色体の偏在につながる可能性があり、不妊症、自然流産、または不均衡な異常を伴う子孫につながる可能性があります。
不均衡な異常は、自発的に、つまり家族歴なしに発生することもあります。子供が不均衡な異常を伴って生きて生まれる可能性は、影響を受ける染色体に大きく依存し、0〜60%の間で変動します。これは、染色体セグメントの喪失(=削除)または重複(=重複)につながります。この文脈では、部分的なモノソミーとトリソミーについても話します。
場合によっては、これらは2つの異なる領域で一緒に発生し、通常、部分的な一染色体が臨床症状の発生に対してより決定的です。これらは削除の顕著な例です 猫の悲鳴症候群とウォルフ・ヒルシュホーン症候群。
光学顕微鏡で変化を判断できなくなったとき、つまり1つまたはいくつかの遺伝子が失われたときの微小欠失について話します。この現象は、プラダーウィリー症候群やアンジェルマン症候群の原因と考えられており、網膜芽細胞腫の発症と密接な関係があります。
ロバートソン転座は特別な場合です:
2つのアクロセントリック染色体(13、14、15、21、22)はセントロメアで結合し、短腕が失われた後に単一の染色体を形成します(構造を参照)。これにより染色体数が減少しますが、これらの染色体の短腕の喪失を簡単に補うことができるため、これはバランスの取れた異常と呼ばれます。ここでも、流産やトリソミーのある生きている子供たちの可能性が非常に高いため、その影響は後の世代でのみ顕著になることがよくあります。
染色体内に2つの切れ目がある場合、中間セグメントが180°回転して染色体に組み込まれる可能性があります。反転として知られるこのプロセスは、ブレークポイントがアクティブな遺伝子(総遺伝物質の2%)内にある場合にのみ不均衡になります。セントロメアが反転セグメントの内側にあるか外側にあるかに応じて、それはペリセントリックまたはパラセントリック反転です。これらの変化はまた、生殖細胞上の遺伝物質の不均一な分布に寄与する可能性があります。
セントロメアが反転セグメントにないパラセントリック反転では、セントロメアが2つまたはまったくない生殖細胞も現れる可能性があります。その結果、対応する染色体は最初の細胞分裂中に失われ、ほぼ確実に流産につながります。
挿入とは、染色体断片を他の場所に設置することです。ここでも、子孫は主に同様の方法で影響を受けます。環状染色体は、特にエンドピースの削除後に発生する可能性があります。シーケンスのタイプとサイズは、症状の重症度を決定します。さらに、これは不正確な分布につながる可能性があり、したがって体細胞内のモザイクタイプにつながる可能性があります。
細胞分裂中に中期染色体が正しく分離しない場合、アイソ染色体が生じる可能性があります。これらは、長い腕または短い腕のみで構成される2つのまったく同じ染色体です。 X染色体の場合、これはウルリッヒ-ターナー症候群(一染色体X)として現れる可能性があります。
このトピックの詳細については、以下をお読みください。 染色体異常
21トリソミー
ダウン症としてよく知られている21トリソミーは、おそらく出生時に最も一般的な数値染色体異常であり、男性の影響がわずかに多くなります(1.3:1)。
21トリソミーが発生する確率は、母親の平均出生年齢などのさまざまな人口統計学的要因に依存し、地域によってわずかに異なります。
21トリソミーの95%は、有糸分裂(生殖細胞分裂)の文脈での分裂エラー、すなわち非分離、すなわち姉妹染色分体の分離の失敗の結果として生じます。
これらは遊離トリソミーとして知られており、母体で90%、父体で5%、胚性ゲノムでさらに5%発生します。
別の3%は、14番染色体または21番染色体のいずれかでの不均衡な乗換えに起因します。 21乗換え、正常染色体と二重染色体21を作成します。残りの2%は、生殖細胞でトリソミーが発生しなかったため、すべての体細胞に影響を与えないモザイクタイプです。モザイクタイプは非常に穏やかであることが多いため、長期間完全に検出されないままになる可能性があります。
いずれにせよ、症状的に同一の遊離トリソミーを、おそらく遺伝性の乗換えトリソミーと区別するために、染色体検査を実施する必要があります。その後、前の世代の家族歴をたどることができます。
このトピックに興味がありますか?これに関する次の記事を読む:21トリソミー
13トリソミー
13トリソミーまたはパトウ症候群の頻度は1:5000で、ダウン症よりもはるかにまれです。原因(無料のトリソミー、乗換え、モザイクの種類)とそれらの割合の分布はほぼ同じです。
理論的には、ほとんどすべての症例は、超音波またはPAPP-Aテストを使用して出生前に診断することができます。 PAPP-A検査は必ずしも定期検査の一部ではないため、中央ヨーロッパの症例の約80%は出生前に診断されます。
超音波検査では、成長の残留物、両側の口唇口蓋裂、異常に小さな目(小眼球症)がすでに見られます。さらに、さまざまな程度の重症度の前脳および顔の奇形が通常存在します(全前脳症)。
大葉型では大脳半球がほぼ完全に分離されて側脳室が作られますが、半葉型では脳の後部だけが分離されて側脳室が失われることがよくあります。最も深刻な形態であるアロバー形態では、大脳半球の分離はありません。
半またはアロバー形状の乳児は通常、出生直後に死亡します。 1か月後の死亡率は出生の約50%です。 5歳まで、13トリソミーからの死亡率は90%に増加します。脳の奇形が原因で、ほとんどの場合、病気の人は一生寝たきりのままで話すことができません。そのため、彼らは完全なケアに依存しています。さらに、Trismoie13の広範囲にわたる身体的症状も存在する可能性があります。
このテーマの詳細については、次を参照してください。 胎児のトリソミー13
16トリソミー
基本的に、16トリソミーは最も一般的なトリソミー(すべてのトリソミーの約32%)ですが、16トリソミーの生きている子供は非常にまれです。一般に、出生は部分的なトリソミーまたはモザイクタイプでのみ発生します。しかし、トリソミーの中で、死産の原因となることが最も多いです。染色体異常による100回の流産のうち32回は、この形態のトリソミーにまでさかのぼることができます。
したがって、主に出生前、すなわち出生前の識別可能な特徴が文書化されています。ここで注目に値するのは、さまざまな心臓の欠陥、成長の鈍化、単一の臍帯動脈(そうでなければ二重)、首の透明度の増加です。これは、リンパ系がまだ完全に発達していないことによる体液の蓄積と、この領域の皮膚の弾力性の増加によって説明されます。さらに、生理的臍ヘルニア、すなわち、腸の大部分がへそを通って外側に一時的に移動することは、しばしば適切に退行せず、これは、臍帯ヘルニアまたは臍帯切断として知られている。
指を交差させた屈曲拘縮も超音波で検出されることがよくあります。少数の出生では、全身性の筋力低下、すなわち全身性の筋力低下が顕著です。これは飲酒の衰弱につながり、乳児に人工的に栄養を与えなければならないことを確実にすることができます。トリソミーの特徴である4本指の溝もよく発生します。ここでも、トリソミーの発生頻度は母親の年齢に直接関係しています。
18トリソミー
エドワーズ症候群、つまり18トリソミーは、1:3000の頻度で発生します。出生前診断では、パトウ症候群と同じです。ここでも、同じ検査ですべての患者を出生前に完全に見つけることができます。原因とその分布は、他のトリソミーと比較できます(21トリソミーを参照)。
さらに、部分的なトリソミーは18トリソミーで発生し、モザイクタイプと同様に、はるかに穏やかな臨床経過につながります。関連する異形性もエドワーズ症候群に非常に特徴的です:出生時、患者の体重は2 kg(通常:2.8-4.2 kg)と大幅に減少し、額は広く後退し、顔の下半分は一般的に発達しておらず、口は小さいです。開口部、狭いまぶたのスリット、後方に回転した、形が変わった耳(額の耳)。新生児のために異常に強く発達した後頭部も目立ちます。肋骨は非常に狭く、壊れやすいです。新生児はまた、筋肉組織全体の永続的な緊張(トーン)を持っていますが、最初の数週間後に生存者で退行します。
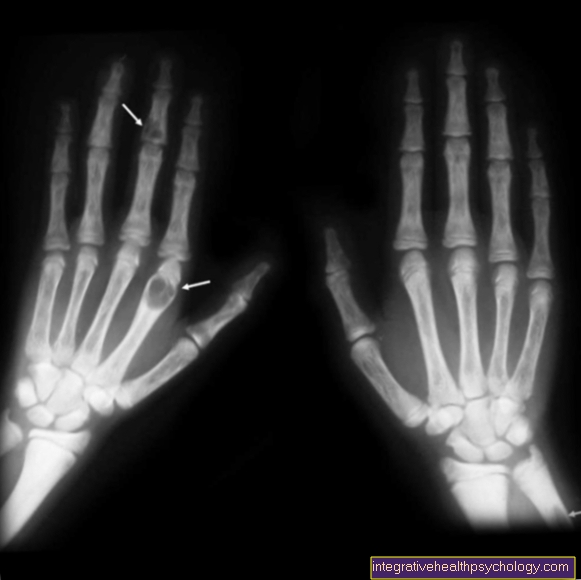
もう1つの特徴は、2番目と5番目の指が3番目と4番目に交差し、指の総数が打ち込まれている一方で、足が異常に長い(経過している)、特に顕著なかかと、発育不全の足指の爪、後退した足の親指があります。 。
深刻な臓器奇形は一般的であり、通常は組み合わせて発生します:心臓と腎臓の欠陥、腸の折り畳み(回転異常)、腹膜の癒着(腸間膜コミューン)、食道の閉塞(食道閉鎖症)など。
これらの奇形のために、死亡率は最初の4日以内に約50%であり、1歳以上で生きているのは約5-10%にすぎません。成人期までの生存は絶対的な例外です。いずれにせよ、知的障害は非常に顕著で話すことができず、寝たきりで失禁しているため、外部の助けに完全に依存しています。
18トリソミーの詳細については、このテーマに関する詳細な記事もお読みください。
- 18トリソミー(エドワーズ症候群)
- 胎児の18トリソミー
トリプルX
トリプルX症候群は、数値染色体異常の最も目立たない形態であり、影響を受ける人々の外観は論理的にすべて女性であり、他の女性と大きく異ならない。特に背が高く、やや「ふっくらとした」顔の特徴があるため、目立つものもあります。精神発達はまた、境界性正常から軽度の精神障害に至るまで、ほぼ正常である可能性があります。
ただし、この知能の不足は、性染色体の他のトリソミー(XXYおよびXYY)よりもいくらか深刻です。 1:1000の頻度では、実際にはそれほどまれではありませんが、トリソミーは通常、臨床的に重大な症状と関連していないため、この病気の女性の大多数は、おそらく生涯にわたって診断されることはありません。
保因者は主に家族の診察中または出生前診断中に偶然発見されます。出産する可能性がわずかに低下し、次世代の性染色体異常の割合がわずかに増加する可能性があるため、子供が欲しい場合は遺伝カウンセリングをお勧めします。
他のトリソミーと同様に、Xトリソミーはほとんどの場合、自由なトリソミーとして発達します。つまり、姉妹染色分体の分裂(非分離)がないためです。ここでも、確率は年齢とともに増加しますが、通常は母体の卵細胞の成熟中に発生します。
脆弱X症候群
脆弱X症候群またはマーティンベル症候群は、X染色体が1つしかないため、変化の影響を受けやすいため、男性に好まれます。
これは、1年間に生きている男性の出生で1:1250の頻度で発生し、非特異的な精神遅滞の最も一般的な形態、つまり、典型的な兆候を伴う特別な症候群では説明できないすべての精神障害になります。
脆弱X症候群は通常、X染色体の1つが偶発的に不活性化されるため、やや弱い形で女の子に発生する可能性があります。スイッチオフされた健康なX染色体の割合が高いほど、症状は強くなります。
しかし、ほとんどの場合、女性は前変異の保因者であり、まだ臨床症状を引き起こしていませんが、息子の完全な突然変異の可能性を大幅に高めています。非常にまれなケースでは、男性が前変異の保因者になることもあり、その場合、通常は臨床的に健康な娘にしか受け継がれません(シャーマンパラドックス)。
この症候群は、FMR遺伝子(脆弱な部位-精神遅滞)のCGGトリプレット(特定の塩基配列)の数が極端に増加することによって引き起こされます。 2000部。
光学顕微鏡下では、これは長い腕の切れ目のように見えます。これが症候群の名前の由来です。これは影響を受けた遺伝子の不活性化につながり、それが今度は症状を引き起こします。
影響を受けた人々は、発話と動きの発達が遅くなり、多動性だけでなく自閉症の方向に進む可能性のある行動上の問題を示す可能性があります。純粋に外部の異常(奇形の兆候)は、目立つあごと耳が突き出た長い顔です。思春期になると、睾丸が大きく大きくなり(マクロオーキディア)、顔の特徴が粗くなります。前変異の女性保因者の間には、心理的異常のわずかな蓄積と特に早期の閉経があります。
染色体分析とは何ですか?
染色体分析は、数値的または構造的な染色体異常を検出できる細胞遺伝学のプロセスです。
このような分析は、たとえば、染色体症候群がすぐに疑われる場合、つまり奇形(異形症)または知的障害(遅滞)の場合だけでなく、不妊症、定期的な流産(流産)の場合にも使用されます。特定の癌(例、リンパ腫または白血病)。
これには通常、患者の血液から得られる特殊なタイプの免疫細胞であるリンパ球が必要です。この方法では比較的少量しか得られないため、細胞を刺激してフィトヘマグルチニンで分裂させ、実験室でリンパ球を培養することができます。
場合によっては、同様の手順で、代わりに皮膚または脊髄からサンプル(生検)を採取します。目的は、現在細胞分裂の真っ只中にあるできるだけ多くのDNA材料を取得することです。中期では、次のステップである後期で細胞の反対側(極)に引き寄せられるように、すべての染色体が細胞のほぼ中央の1つのレベルに配置されます。
この時点で、染色体は特に密集しています(高度に凝縮されています)。紡錘体毒コルヒチンが追加されます。これは細胞周期のこの段階で正確に機能するため、中期染色体が蓄積します。次に、それらを分離し、特別な染色方法を使用して染色します。
最も一般的なのはGTGバンディングで、染色体はトリプシン、消化酵素、および色素ギムザで処理されます。特に密集した地域とアデニンとチミンが豊富な地域は暗く示されています。
得られたGバンドは各染色体に特徴的であり、簡単に言えば、遺伝子の少ない領域と見なされます。このように染色された染色体の写真は1000倍の倍率で撮影され、コンピュータープログラムの助けを借りて核型が作成されます。バンドパターンに加えて、染色体のサイズとセントロメアの位置を使用して、それに応じて染色体を配置します。しかし、非常に異なる利点を持つことができるバンディングの他の方法もあります。
編集チームからの推薦
より一般的な情報については、次の記事を参照してください。
- 細胞核分裂
- 細胞核の機能
- 21トリソミー
- 遺伝病






.jpg)